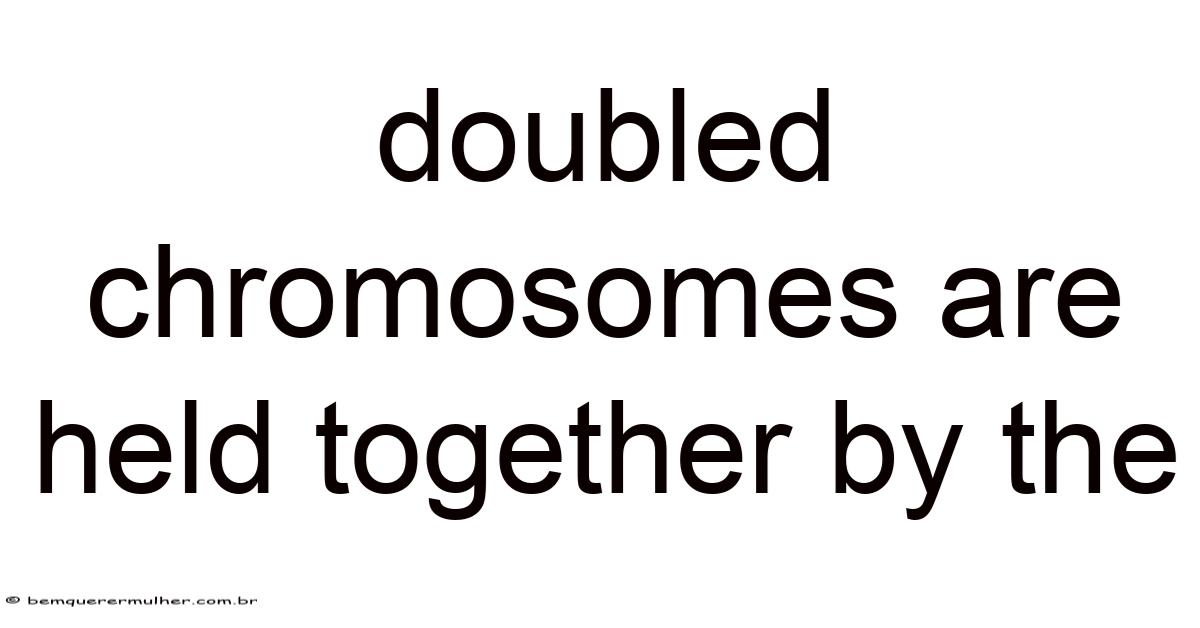
Doubled Chromosomes Are Held Together by Cohesin Complexes
Doubled chromosomes, scientifically known as sister chromatids, are the identical copies formed when a chromosome undergoes DNA replication during the S phase of the cell cycle. So naturally, these duplicated chromosomes must remain precisely held together until the appropriate moment during cell division. The molecular machinery responsible for this crucial task is the cohesin complex, a ring-shaped protein structure that embraces sister chromatids like a molecular embrace, ensuring their proper segregation during cell division Took long enough..
Understanding Chromosome Structure and Replication
Before delving into the mechanisms of sister chromatid cohesion, it's essential to understand chromosome structure and replication. Chromosomes are composed of DNA molecules tightly coiled around histone proteins, forming chromatin. During the S phase of the cell cycle, each chromosome is replicated, resulting in two identical DNA molecules known as sister chromatids. These chromatids remain connected along their entire length until anaphase, the stage of mitosis or meiosis when they separate and move to opposite poles of the dividing cell.
Honestly, this part trips people up more than it should And that's really what it comes down to..
The connection between sister chromatids is not merely a passive association but an active, regulated process critical for accurate chromosome segregation. Without proper cohesion, sister chromatids would separate prematurely, leading to aneuploidy—a condition where daughter cells have an incorrect number of chromosomes, which can result in cell death or diseases like cancer.
The Cohesin Complex: Molecular Glue of Chromosomes
The primary structure responsible for holding sister chromatids together is the cohesin complex. This multi-subunit protein complex forms a ring-like structure that topologically encircles the sister chromatids, effectively linking them together. The cohesin complex consists of several protein subunits, including:
- SMC1 and SMC3 (Structural Maintenance of Chromosomes proteins): These form the ATPase-containing "head" and "hinge" domains of the cohesin ring
- RAD21 (also known as SCC1 or Mcd1 in yeast): This subunit bridges the SMC1 and SMC3 proteins, completing the ring structure
- SA1/SA2 (also known as STAG1/STAG2): These regulatory subunits help in cohesin loading and stabilization
The ring structure of cohesin is thought to trap sister chromatids within its central cavity, preventing their separation until the proper signal for anaphase onset is received. This "ring model" suggests that cohesin holds sister chromatids together by encircling both DNA molecules simultaneously.
Loading and Establishment of Cohesin
Cohesin loading onto chromosomes occurs during the G1 phase of the cell cycle, before DNA replication begins. This process is facilitated by a separate loading complex that includes the Scc2-Scc4 (NIPBL-MAU2 in humans) proteins. Once loaded onto chromosomes, cohesin complexes distribute along the chromatin arms.
During DNA replication in the S phase, the replication machinery passes through the cohesin rings. This process establishes the connection between newly replicated sister chromatids. Interestingly, a significant portion of cohesin is displaced during replication, with new cohesin complexes being loaded afterward to ensure proper cohesion Turns out it matters..
It's the bit that actually matters in practice.
Regulation of Cohesin Removal
The precise timing of sister chromatid separation is critical for accurate chromosome segregation. Cohesin removal is tightly regulated and occurs in two distinct phases:
-
Cleavage-dependent removal: During prophase and prometaphase of mitosis, a subset of cohesin complexes is removed from chromosome arms by a process called "prophase pathway." This removal is mediated by WAPL (Wings apart-like protein), which opens the cohesin ring and allows it to slide off the DNA.
-
Cleavage of cohesin subunits: The majority of cohesin complexes remain on chromosomes until anaphase onset. At this stage, the protease separase cleaves the RAD21 subunit of cohesin, irreversibly breaking the cohesin ring and allowing sister chromatid separation Worth keeping that in mind. Practical, not theoretical..
The activation of separase is itself tightly regulated by securin, a protein that inhibits separase until the proper time. At anaphase onset, securin is degraded via the anaphase-promoting complex/cyclosome (APC/C), releasing separase to cleave RAD21.
Importance of Proper Sister Chromatid Cohesion
Proper sister chromatid cohesion is essential for accurate chromosome segregation during cell division. When cohesion functions correctly:
- Sister chromatids align properly at the metaphase plate
- Chromosomes experience correct tension at the kinetochore during mitosis
- Segregation errors are minimized
Defects in sister chromatid cohesion can lead to severe consequences:
- Premature sister chromatid separation: Results in unequal chromosome distribution
- Chromosome mis-segregation: Leads to aneuploidy in daughter cells
- Genomic instability: Associated with developmental disorders, infertility, and cancer
Cohesion in Meiosis
While the basic principles of cohesin function are conserved between mitosis and meiosis, meiosis involves additional complexity. During meiosis, cohesin plays specialized roles in:
- Homologous chromosome pairing and synapsis
- Ensuring proper recombination between homologous chromosomes
- Maintaining cohesion at centromeres until anaphase II, while arm cohesion is released at anaphase I
These specialized functions are crucial for proper chromosome segregation in meiosis and the production of haploid gametes.
Cohesin and Human Disease
Given the fundamental importance of sister chromatid cohesion, it's not surprising that defects in cohesin function or regulation are associated with human diseases:
- Cohesinopathies: A group of developmental disorders caused by mutations in cohesin subunits or regulatory proteins, including Cornelia de Lange syndrome and Roberts syndrome
- Cancer: Aberrant cohesin function is observed in various cancers, contributing to chromosomal instability
- Infertility: Defects in meiotic cohesion can lead to infertility due to failed gamete production
Research Advances and Future Directions
Research on sister chromatid cohesion has advanced significantly in recent years, with new insights into:
- The three-dimensional organization of chromosomes mediated by cohesin
- The role of cohesin in gene regulation and chromosome architecture
- The mechanisms of cohesin loading and unloading
- The relationship between cohesin and other cellular processes
Future research aims to further elucidate the molecular details of cohesin function and develop therapeutic strategies for diseases associated with cohesion defects.
Conclusion
Doubled chromosomes are held together by the cohesin complex, a remarkable molecular machine that ensures the faithful segregation of genetic material during cell division. This nuanced system of sister chromatid cohesion is essential for maintaining genomic stability and preventing diseases associated with chromosomal abnormalities. Worth adding: as our understanding of cohesin continues to deepen, we gain not only fundamental insights into chromosome biology but also potential avenues for treating diseases caused by cohesion defects. The molecular embrace of cohesin represents one of nature's most elegant solutions to the challenge of accurately duplicating and distributing our genetic blueprint It's one of those things that adds up..
Translating Cohesin Biology into Therapeutic Opportunities
The mechanistic insights gained over the past decade have begun to shape how researchers approach disease treatment. Small‑molecule inhibitors that disrupt the interaction between cohesin and its loader, SCC‑1 (RAD21), have shown promise in sensitizing cancer cells that are dependent on elevated cohesin activity for survival. Notably, tumors harboring mutations in the STAG2 or SMC1A genes often display a “cohesin‑addiction” phenotype, making them vulnerable to pharmacological interference with the ATPase domain of SMC1/SMC3. Early‑stage preclinical studies using ATP‑competitive inhibitors have demonstrated selective cytotoxicity in cell lines and patient‑derived organoids, raising the prospect of precision oncology strategies that exploit cohesion defects rather than simply correcting them.
Short version: it depends. Long version — keep reading.
Beyond oncology, the emerging link between cohesin and epigenetic regulation has opened a new avenue for treating neurodevelopmental disorders. Recent CRISPR‑based screens in induced pluripotent stem cell (iPSC)‑derived neural progenitors have identified compounds that restore proper enhancer looping and rescue transcriptional programs, suggesting that modulating chromatin architecture could ameliorate developmental phenotypes. Mutations in the NIPBL gene, which encodes the primary loader of cohesin onto chromatin, not only cause Cornelia de Lange syndrome but also alter the distribution of histone marks and enhancer‑promoter contacts that are critical for neuronal differentiation. Such approaches underscore the broader principle that cohesion defects are not merely structural failures but also regulatory missteps that can be corrected through targeted biochemical interventions Most people skip this — try not to..
Technological Frontiers: Visualizing Cohesin in Action The past five years have witnessed a revolution in how scientists observe cohesin dynamics within living cells. Super‑resolution microscopy techniques such as single‑molecule localization microscopy (SMLM) and lattice light‑sheet imaging now permit the tracking of individual cohesin complexes over time, revealing stochastic loading and unloading cycles that were previously invisible. Also worth noting, proximity‑labeling methods—including auxin‑induced degron tagging coupled with mass spectrometry—have enabled researchers to map the interactome of cohesin in a cell‑type‑specific manner, uncovering previously unknown partners such as the chromatin remodeler CHD4 and the transcription factor CTCF‑associated co‑activators. These high‑resolution tools are reshaping the conceptual framework of cohesion, moving it from a static “glue” model to a dynamic, regulated hub that integrates signals from DNA replication, transcription, and DNA damage response pathways.
Synthesis: From Molecular Embrace to Systemic Implications
Taken together, the advances described above illustrate how the once‑simple notion of sister chromatid cohesion has expanded into a multifaceted discipline that bridges basic cell biology, disease genetics, and therapeutic development. The cohesin complex is now recognized not only as a custodian of chromosome segregation but also as a central orchestrator of three‑dimensional genome organization, gene regulation, and cellular homeostasis. Its dysfunction reverberates across a spectrum of pathologies—from developmental syndromes to malignancies—making it an attractive focal point for interdisciplinary research.
Conclusion
Understanding how doubled chromosomes are tethered together by the cohesin complex offers more than a glimpse into the mechanics of cell division; it provides a foundational framework for interpreting the structural and regulatory underpinnings of genomic stability. As researchers continue to decode the molecular choreography of cohesin loading, unloading, and functional interaction with other chromatin regulators, the insights gained will increasingly inform strategies to correct cohesion defects at their root. Whether through the development of small‑molecule modulators, gene‑editing‑based rescue of mutated loader proteins, or the engineering of synthetic cohesion circuits for synthetic biology applications, the future holds promise for translating the elegant molecular embrace of cohesin into tangible health benefits. In this way, the study of sister chromatid cohesion stands as a testament to how fundamental biological principles can be leveraged to address some of humanity’s most pressing medical challenges.